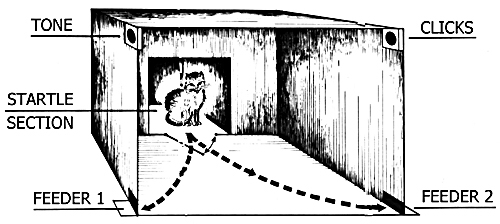
The choice of learning task. While discussing the significance of hippocampus in the organization of learning and memory special attention is devoted to the type of the tasks to be used. Memory is classified mainly as the hippocampal-dependent and non-dependent types (O'Keefe and Nadel 1978, Nadel 1991, 1995, Mooris et al. 1982, 1990, Squire 1992, Zola-Morgan et al. 1986, Zola-Morgan and Squire 1993, Rempehl-Clower et al. 1996). Particularly sensitive to the intactness of the hippocampus is the so-called "spatial memory" which is assigned to the hippocampal-dependent type (Olton et al. 1978, Olton 1979). The task used in our experiments, being rather complex, falls into this category. Cats were required to remember the location of food which, in response to one sound stimulus (tone) was delivered from the left feeder, while in response to the other (clicks), from the right. As the appearance of these signals was randomized and their location was not altered, animals permanently were posed with a task to go to the right or to the left. Thus the problem posed before the animals was an example of spatial instrumental sound discrimination and, proceeding from the above-said classification, had to be sensitive to the RFI of hippocampus.
The choice of a method for RFI of hippocampus. It is known that the electrolytic lesion or surgical removal of any brain structures leads to the irreversible structural changes and compensatory plastic developments in the synaptic terminals and all of this in the structures of limbic circuit is realized more easily. Therefore in order to the understanding of functional significance of any brain structures the method of RFI is considered to be more adequate. Different times to the aim of RFI a various methodical approach were used: ECS (McGaugh and Zornetzer 1970, McIntire and Molino 1972), local electrical stimulation of brain structures (Kesner and Doty 1968, Vardaris and Schwartz 1971, Kapp et.al. 1974, Kesner 1979), focal (Brooks 1983) and total (Hundwerker et al. 1975) cooling, injection of agonists and antagonists of various neurotransmitters (Martin 1991, Jerusalinsky et al. 1992), spreading depression (Bures et al. 1974), injection of local anesthetics (Albert and Madriga 1980, Sandkuchler et.al.1995, Bohbolt et al. 1996), focal injection of tetrodotoxin (Cahil et al. 1987, Zhuravin and Bures 1991) etc. A detailed analysis of all the abovementioned methods of RFI indicates, that many of those, has a definite limitations also. Particularly, the use of focal and total cooling, injection of mediator system agonists and antagonists, spreading depression or injection of anesthetics in the studies of memory mechanisms and time parameters of consolidation is appropriate in the case of one trial learning only. In many- trial and many- session learning (elaboration of active avoidance, learning in mazes, elaboration of discrimination of conditioned signals etc.) given methods becomes inadequate. In this respect, the use of threshold electrical stimulation, inducing local epileptiform discharges in a definite brain structures, seems more appropriate.
Effect of RFI of hippocampus on the motivational-emotional state of animals. When the method of induction of local epileptiform discharges was used for the aim of RFI of hippocampus, special attention was paid to the animals' motivational-emotional state. No considerable change of this parameter was observed in our experiments. This fact has an important methodical significance. As the matters of fact, as been mentioned above, in the early works of authors (Leonard and Zavala 1964, Hurdspeth et al. 1964) it was indicated that the ECS, applied prior to and after learning trials, alter the physical state of animals. They show fear before the experimental set, refuse to enter the experimental cabin. Though the experiments of those authors essentially differed from ours, in our case the evoked epileptiform discharges were restricted only to the hippocampus not involving other brain structures in pathological activity, but in our experiments too they were elicited after each combination of conditioned and unconditioned stimuli. In spite of this, development of fear reaction was not observable, animals without any enforcement approached that feeder at which had earlier received electrical stimulation. This fact leads to the conclusion that the local epileptiform discharges, elicited in the hippocampus several times during one learning session, do not change significantly the animals' general behavior. Similar result once more emphasizes the advantage of application of the method of local electrical stimulations of definite brain structures over the ECS.
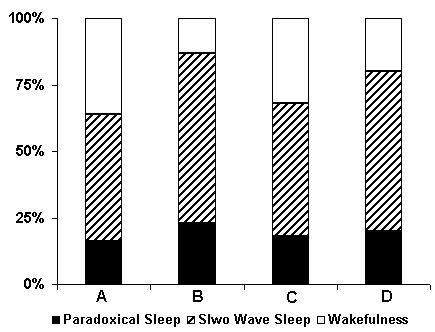
Influence of hippocampal RFI with various intervals after each pairing of conditioned and unconditioned signals on SWC. In our experiments during studding of influence of RFI of hippocampus after each pairing of conditioned and unconditioned signals in the course of elaboration of spatial sound discrimination with food reward, an interesting fact was shown - a serious changes of structure and percentage of SWC; sharp enhancement of slow wave and paradoxical phases of sleep. The mechanism of indicated changes is very difficult for explanation, that is why the SWC undergoes simultaneously to two kinds of influences in these cases: 1) to the processes of learning and 2) to the epileptiform activity.
There is a controversy in the literature about the problem of influence of learning processes on the subsequent SWC. Some authors noted an increase of paradoxical sleep after learning session (Fishbein and Gutwein 1977), others, indicate to the absence of any noticeable effect (Oniani and Lortkipanidze 1985). Our results exclude the existanse of a causal relationship between learning processes and postsessional enhancement of paradoxical sleep amount.
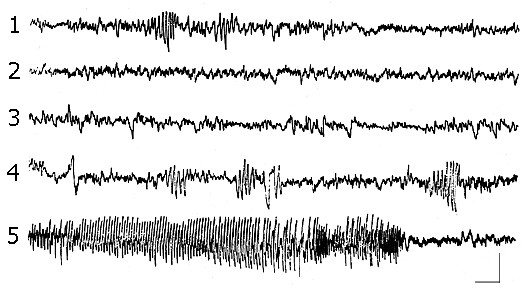
Some kind of disagreement exists in the experimental evidence concerning the influence of epileptiform activity on the SWC. Here is underlined a strong dependence of a character of effects on the expression of epileptiform discharges (Mutani 1967, Roldan 1969). There was used a threshold electrical stimulation in our experiments, leading to the development of EEG epileptiform discharges, which were located in the hippocampus. Those cases, when the epileptiform discharges occupied the electrical activity of other brain structures and/or manifested behaviorally, were rejected.
Evidence concerning the similarities of SWC changes both under the influence of only hippocampal repetitive epileptiform discharges and after the impact of RFI of this structure after each pairing of conditioned and unconditioned signals, rules out once again the possibility about causal relationship of these changes with the consolidation of short term memory traces during conditioning. We believe that, repititive epileptiform discharges, originating in hippocampus, through their descending influence leads to the strong activation of mesodiencephalic structures, triggering SWS, and as a result of later there occurs a significant increase of paradoxical sleep.
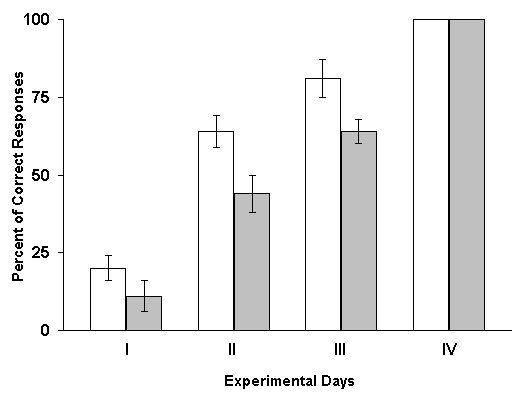
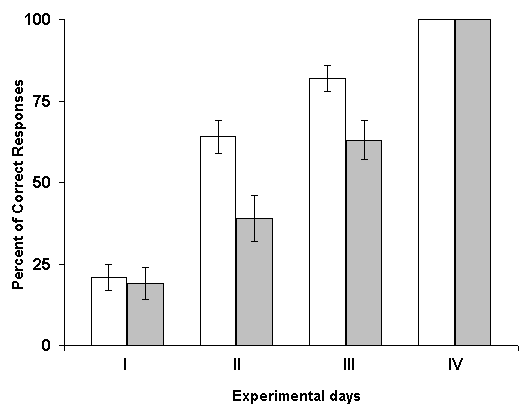
Influence of RFI of hippocampus on the elaboration of instrumental spatial sound discrimination. Experiments with the study of the effects of RFI by means of induction of local epileptiform discharges after daily sessions of elaboration have demonstrated that, the animals of experimental group somewhat lag behind the controls by percentage of correct responses at the beginning learning sessions. Despite of this, a 100% level of discrimination was achieved on the fourth session for both groups. If the normal functioning of the hippocampus actually is necessary for the consolidation and formation of memory traces, its RFI in the initial phases of learning should then have exerted especially powerful impairing influence, because the RFI of this structure occupied namely that period, during which the process of consolidation must be realized most intensively. Indeed, in our experiments it dos not so; the processes of consolidation of memory traces was accomplished in experimental animals with same rates as in controls. Similar results were obtained also with the induction of hippocampal RFI prior to the initiation of everyday learning sessions. Analyzing the data of this series of experiments it could be assumed that the critical factor contributing to the normal course of acquisition of spatial sound discrimination in these experiments is that in the periods between the paring of conditioned and unconditioned stimuli the functional state of the hippocampus remains normal and, consequently, it can participate in the consolidation of memory traces. In order to check this possibility in the next series of experiments electrical stimulation of the hippocampus, eliciting its RFI, was applied in the periods between the combination of conditioned and unconditioned stimuli.
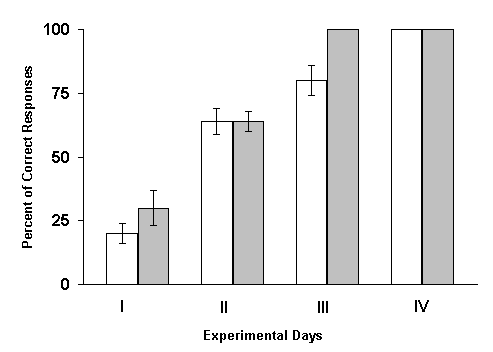
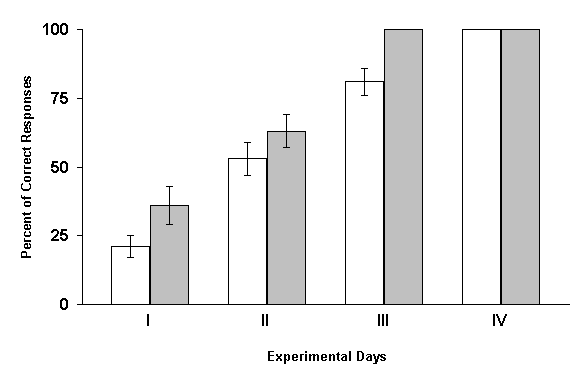
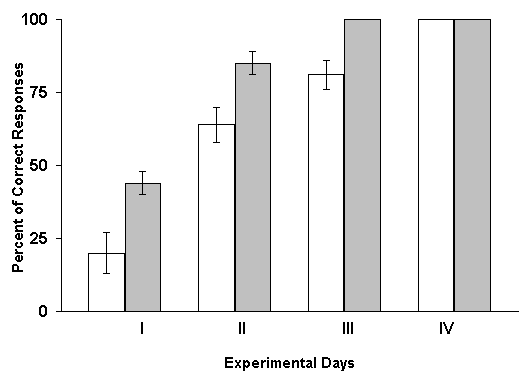
The results of these experiments turned out to be diametrically opposite. Under these conditions one could observe not the delay of learning, but on the contrary, its facilitation. At RFI of the hippocampus, with 1 min intervals after every paring of conditioned and unconditioned stimuli, elaboration of spatial sound discrimination with food reward was completed in experimental animals a day earlier than in controls. Analysis of these data lend support to the conjecture that one minute during which after each paring the hippocampus remains without the intervention of local epileptiform discharges appears to be sufficient for this structure to participate in the fixation of memory traces. Therefore, in the subsequent series of experiments the interval between the combinations of conditioned and unconditioned stimuli and elicitation of hippocampal RFI was still more shortened. It appeared that, in these cases, the facilitating influence of RFI, become more pronounced. The most significant facilitation of elaboration of the spatial sound discrimination with food reward was observed when the hippocampal RFI was made just immediately after each paring of conditioned and unconditioned stimuli. Similar results make one think that either consolidation of traces of spatial memory, studied in our experiments, does not realized in the hippocampus, or the processes of fixation of this information are too short lasting and are over before the interference of epileptiform discharges. Let us try to analyze how much this conclusion is in conformity with the data reported in the literature. As been indicated above, the effects of electrical stimulation of the hippocampus applied after learning trials are rather controversial. It is difficult to interpret the discrepancy in the results because of variation of learning situations, the tasks to be learned, difference in the stimulation conditions, etc. However, we shall still try to make some remarks. In the studies of some authors (Brunner et al. 1970, Lidsky and Slotnick 1970, 1971, Haycock et al. 1973, Kesner and Conner 1973, Sideroff et al. 1974) it has been demonstrated that the sub-threshold electrical stimulation of the hippocampus brings about the impairment of task learning. All these data become difficult to be explained also for that reason that with the application of such parameters of stimulation of the hippocampus which do not cause convulsive activity in this structure, many authors obtained facilitation of learning of a variety of tasks (Landfield et al. 1973, Destrade et al. 1973, Soumireu-Mourat et al. 1975, Destrade and Jaffard 1978).
While discussing the effects of threshold electrical stimulation of the hippocampus causing development of epileptiform discharges, critical becomes the question about the sphere of extension of convulsive activity. Clarification of this question is of great importance, since in the works of a number of authors it was supposed that the reason for memory disturbance is the spread over other brain structures of convulsive activity originating in the hippocampus (Kesner and Doty 1968, Vardaris and Schwartz 1971, Kapp et al. 1974, Kesner 1979). On the basis of all of these it becomes evident that in the experiments, aimed in a studding of the effects of RFI of any brain structure by means of its' electrical stimulation, a precise electrophysiological control is a necessary one condition. Such a supposition is supported by the results of many studies, indicating on the lack of effects on the learning processes in the case of electrical stimulation of hippocampus, eliciting local epileptiform discharges in it (Hirano 1966, Kesner and Doty 1968, Vardaris and Schwartz 1971, Kapp et al. 1974).
The data on the facilitating influence of epileptiform discharges on the process of elaboration of spatial sound discrimination with food reward, obtained in our experiments, are in keeping with those obtained by some authors earlier (Chow 1961, Stein and Chorover 1968, Erikson and Patel 1969). In the experiments of Gralewitz (1976) electrical stimulation of the hippocampus, not eliciting epileptiform discharges, led to the facilitation of learning, while more powerful stimulations, leading to the initiation of epileptic activity, on the contrary facilitate the learning. However challenging is the fact that in these experiments the conditioned signal and electrical stimulation of the hippocampus were delivered simultaneously. Under such experimental conditions the occurrence of the hippocampal epileptiform discharges has a principal significance, since it has been demonstrated that (Nachkebia and Oniani un-publicated data) the conditioned signals are perceived by the animals neither against the background of hippocampal epileptiform discharges, nor during a certain time after their cessation. Naturally it can be supposed that in these conditions learning might be hindered or blocked because of the absence of perception of conditioned signals and not for other reasons.
Yet, the facilitating influence of RFI of the hippocampus is in need of exhaustive explanation that has not been done in those works. Useful information on the reasons for learning facilitation under the influence local hippocampal epileptiform discharges, elicited by electrical stimulation after each combination of conditioned and unconditioned signals can be provided at a parallel observation made on the dynamics of the neocortical electrical activity. In the investigations of Oniani and Nachkebia (1982) it has been shown, that against the local hippocampal epileptiform discharges electrical activity of the neocorotex undergoes a diffuse desynchronization as is the case in response to the stimulation of the mesodiencephalic activating structures. This phenomenon was permanently observed in the above indicated experiments. This fact indicates that when epileptiform discharges are initiated in the hippocampus, the mesodiencephalic unspecific activating structures get involved in excitation, which in their turn, have an activating influence on the neocortex, thereby creating an optimal background for the receipt and recording of information. Moreover, the facilitation of the receipt and recording of information by the brain is so much vigorous that it may supposedly not only mask the negative influence of RFI of the hippocampus and disturbance of the normal functioning of the septo-hippocampal system, but also creates the background for the facilitation of learning just in the initial stages of acquisition of a variety of tasks, when the receipt and recording of new information proceed most intensively. Similar interpretation is in keeping with the theory of Hebb (1949) concerning the significance of the level of reticular activation for information processing in the brain. In this aspect noteworthy are the data of Bloch and Laroche (1985), who have shown that electrical stimulation of MRF, causing improvement of learning, enhances the long-term potentiation in the hippocampus. At the same time, the induction of the facilitating influence appeared to depend on the duration of interval between the trial and the onset of MRF electrical stimulation. The most remarkable enhancement of long-term potentiation was obtained at 10 sec interval, while the lengthening of this interval reduced the facilitating influence. Results, suggesting the relation of the long-term potentiation with learning, were obtained by other authors (McNaughton et al. 1986, Davis et al. 1992) as well.
If one assumes that in the learning process the hippocampus serves the function of transferring of information for recording then, from this point of view, the facilitating influence of its RFI may be accounted for more fully. Usually, during the elaboration of any task, animals receive a mass of information except that which is the primary at that time. When RFI of hippocampus appears following different intervals after each paring of conditioned and unconditioned stimuli, the amount of information, arriving in the hippocampus, is restricted. The level of restriction varies depending on the interval passed between the reinforcement of conditioned signal with food and the electrical stimulation of the hippocampus, causing RFI of this structure. This is indicated by the fact that at the shortening of this interval the facilitating influence becomes more pronounced, achieving its maximum at the zero interval, when RFI of the hippocampus occurs immediately after the transferring of basic information for recording and, during a definite time, access to the hippocampus or transfer from it of any information is ruled out. This is supplemented also by excitation of mesodiencephalic unspecific activating structures during development in the hippocampus of epileptiform discharges, thereby creating optimal background for information recording and storage in long-term memory. The long-term potentiation was not studied in our experiments however the operation of the same mechanism described by Bloch and Laroche (1985) can be speculated in this case as well. As to the case of RFI of the hippocampus prior to and after daily learning sessions, the interval between the separate trials and the onset of hippocampal electrical stimulation, eliciting epileptiform discharges, is too long for this influence to be exerted.
The suggestion that the hippocampus may serve the function of transferring of the information for recording is not by itself new. It takes a central position in the theory of Vinogradova (1975). According to this theory, the hippocampus transfers information for recording after determining of novelty of the arriving signals. In our point of view, recorded is the information, which is of definite importance for the organism. For the triggering of the mechanisms for information recording, particular significance must have the informational charachteristics of the stimulus, particularly, the degree of the stimulus elicited motivational-emotional reactions, the electrographic correlates of which is the development of the hippocampal theta rhythm (Oniani and Badridze 1974). If one takes into account the views of some authors (Vinogradova 1975, Rudell et al. 1980, Gray 1982), that the function of theta rhythm is the selection of the stimuli, arriving in the hippocampus, it may be then suggested, that the process of selection starts with the determination of their value after which the signals of a definite importance to the organism are transferred by the hippocampus to be recorded for the long-term memory storage.
Albert, D.J. and Madryga, F.G. An examination of the functionally effective spread of 4 ml of slowly infused lidokaine. Behavioral and Neur. Biol., 1980, 29: 378-384.
Ambrogi Lorenzini, C.G., Baldi, E., Bucherelli, C., Sacchetti, B. and Tassoni, G. Neural topography and chronology of memory consolidation: A review of functional inactivation findings. Neurobiol. of Learning and Memory. 1999, 71: 1-18.
Barcik, J.D. Hippocampal afterdischarges and memory disruption. Proc. Amer. Psychol. Ass., 1969, 4: 185-186.
Barcik, J.D. Hippocampal afterdischarges and conditioned emotional response. Psychol. Sci. 1970, 20: 297-299.
Bloch, V. and Laroche S. Enhancement of long term potentiation in the rat dentate gyrus by post-trial stimulation of the reticular formation. J. of Physiol., 1984, 360: 215-231.
Bohbot, V., Otahal, P., Liu, Z., Nadel, L. and Bures, J. Electroconvulsive shock and lidocaine reveal rapid consolidation of spatial working memory in the water maze. Proc. of the Nat. Acad. of Sci. of USA, 1996, 93: 4016-4019.
Brooks, V.B. Study of brain function by local, reversible cooling.Rev. of Physiol., Biochem. and Pharmacol., 1983, 95: 1-109.
Brunner, R.L., Rossi, R.R., Sturz, R.M. and Roth T.G. Memory loss following post trial electrical stimulation of the hippocampus. J. of Psych. Sci., 1970, 18: 159-160.
Bures, J., Buresova, O. and Krivanek, J. The mechanisms and applications of Leao's spreading depression of electroencephalographic ativity. 1974, New York: Academic Press.
Cahil, L., Coopersmith, R.M., Leon, M. and McGaugh, J.L. Local injection of tetrodotoxin decreases metabolic activity in discrete brain regions. A 2-deoxiglucose autoradiography analysis. Soc. of Neurosci. abst., 1987, 13: 1414.
Chow, K.L. Effects of local electrographic after discharges on visual learning and retention in monkey. J. of Neurophysiol., 1961, 24: 391-400.
Davis, S., Butcher, S.P. and Morris, R.G.M. The NMDA receptor antagonist D-2-amino-5 phosphonopentanoate (DAP-5) impairs spatial learning and LTP in vivo at intracerebral concentrations comparable to those that block LTP in vitro. J. Neurosci. 1992, 12: 21-34.
Destrade, C. and Jaffard, R. Post-trial hippocampal and lateral hypothalamic electrical stimulation: facilitation of long-term memory of appetitive and avoidance learning tasks. J. of Behav. Biol., 1978, 22: 354-374.
Destrade, C., Soumireu-Mourat, B. and Cardo, B. Effects of post-trial hippocampal stimulation on the acquisition of operant behavior in the mouse. J. of Behav. Biol., 1973, 8: 713-724.
Douglas, R.J. The hippocampus and behavior. Psychol. Bull., 1967, 67: 416-442.
Erickson, C.K. and Patel, J.B. Facilitation of avoidance learning by post-trial hippocampal electrical stimulation. J. of Comp. Physiol. and Psychol., 1968, 68: 400-406.
Fernandez, G., Weyerts, H., Schrader-Bolsche, M., Tendolkar, I.,Smid H.G., Tempelmann, C., Hinrichs H., Scheich, H., Elger, C.E., Mangun, G.R. and Heinze H.J. Successful verbal encoding into episodic memory engages the posterior hippocampus: a parametrically analyzed functional magnetic resonance imaging study. J. of Neurosci., 1998, 18: 1841-1847.
Fishbein, W. and Gutwein, M. Paradoxical sleep and memory storage processes. Behav. Biol., 1977, 19, 3: 425-464.
Gaffan, D. Recognition impaired and association intact in the memory of monkeys after transection of fornix. J. of Comp. Physiol. and Psychol., 1974, 86: 1100-1109.
Gaffan, D. Monkeys' recognition memory for complex pictures and the effect of fornix transection. J. of Exp. Psychol., 1977, 29: 505-514.
Gluck, M.A. and Mayers C.E. Hippocampal mediation of stimulus representation: A computational theory. Hippocampus, 1993, 4: 491-516.
Gralewitz, S. Electrical stimulation of the hippocampus and acquisition of the conditioned avoidance response in shuttle-box in cats. Acta Neurobiol. Exp., 1976, 36: 640-654.
Gray, J. A, The Neuropsychology of Anxiety: an Enquiry into the Functions of the Septo-Hippocampal System. Clarendon Press, Oxford, Oxford University Press, New York, 1982.
Greenberg, R., Pearlman, C., Brooks, R., Mayer, R. and Hartmann, E. Dreaming and Korsakoff's psychosis. Arch. of Centr. Psych., 1968, 18: 203-209.
Haycock, J.W., Deadwyler, S.A., Sideroff, G.I. and McGaugh, J. L. Retrograde amnesia and cholinergic systems in the caudate-putamen complex and dorsal hippocampus of the rat. Exp. Neurol., 1973, 41: 201-213.
Hebb, D.O. The Organization of Behavior. Wiley, New York, 1949.
Hirano, T. Effect of hippocampal electrical stimulation on memory consolidation. Psychol., 1966, 9: 63-75
Hundspeth, W.J., McGaugh J.L., Thompson C.W. Aversive and amnesic effects of electroconvulsive shock. J. of Comp. Physiol. and Psychol., 1964, 57: 61-64.
Handwerker, H.O., Iggo, A. and Zimmerman M. Segmental and supraspinal action on dorsal horn neurons responding to noxious and non-noxious skin stimuli. Pain 1, 1975: 145-165.
Isaacson, R.L. The Limbic System. Plenum Press, New York-London, 1976.
Jasper, H.H., Ajmone-Marsan C. A Stereotaxic Atlas of the Diencephalons of the Cat. Ottawa Nat. Res. Council, Canada, 1954.
Jerusalinsky, D., Ferreira, M.B.C., Walz, R., Da Silva, R.C., Bianchin, M., Ruschel, A., Median, J.H. and Izquierdo, I. Amnesia by infusion of glutamate receptor blockers into the amigdala, hippocampus and entorhinal cortex. Behav. and Neur. Biol., 1992, 58: 76-80.
Kapp, B.S., Kaufman, G.D., Repole, D.A. Hippocampal stimulation and memory: effects of stimulation parameters and reinforcement magnitude. Physiol. and Behav., 1974, 13: 47-55.
Kesner, R.P. Hippocampus and memory. Behav. Brain Sci., 1979, 2: 413-437.
Kesner, R.P. and Conner H.S. Indipendence of short and long-term memory: a neural system analysis. Science, 1973, 176: 432-434.
Kesner, R.P. and Doty, R.W. Amnesia produced in rats by local seizure activity initiated from the amigdala. Exp. Neurol., 1968, 21, 1: 58-68.
Kim, C., Choi, H., Kim, C.G., Kim, J., Kim, M.S., Park, H.L., Ahn, B.T. Effect of hippocampectomy on sleep pattern in cats. Electroenceph. Clin. Neurophysiol., 1975, 38: 235- 243.
Kimble, D.P. Hippocampus and internal inhibition. Psychol. Bull., 1968, 70: 285-295.
Kluver, H. and Bucy, P.C. Preliminary analysis of the temporal lobes in monkeys. Arch. of Neurol. and Psych., 1939, 42: 979-1000.
Landfield, P.W., Tusa R.J. and McGaugh J.L. Effects of posttrial hippocampal stimulation on memory storage and EEG activity. Behav. Biol., 1973, 8: 485-505.
Leonard, D.J. and Zavala, A. Electroconvulsive shock, retrograde amnesia and the single-shock method. Science, 1964, 146: 1073-1076.
Lidsky, A. and Slotnick, B.M. Electrical stimulation of the hippocampus and electroconvulsiveh shock produce similar effects in mice. Neuropsychol., 1970, 8: 363-369.
Lidsky, A. and Slotnick, B.M. Effects of post-trial limbic stimulation on retention of a one-trial passive avoidance response. J. Comp. Physiol. Psychol., 1971, 76: 337-348.
Martin, J.H. Autoradiographic estimation of the extent of reversible inactivation produced by microinjection of lidocaine and muscimol in the rat. Neurosci. Lett., 1991, 127: 160-164.
McLean, P.D. Psychosomatic disease and the "visceral brain". Psychosom. Med., 1949, 11: 338-353.
McDonough, I.H. and Kesner, R.P. Amnesia produced by electrical stimulation of amygdale and dorsal hippocampus in cats. J. Comp. Physiol. Psychol. 1971, 77: 171-178.
McGaugh, J.L. and Zornetzer, S.E. Amnesia and brain seizure activity in mice: effects of diethile anesthesia prior to electroconvulsive shock stimulation. Behav. Biol., 1970, 5: 243-248.
McIntire, C. and Molino A. Amigdala lesions and learning: long-term effect of kindling. Physiol. and Behav. 1972, 8: 1055-1062.
McNaughton, B.L., Barnes, C.A., Rao, G., Baldwin, J. and Rasmussen, M. Long-term enhancement of hippocampal synaptic transmission and the acquisition of spatial information. J. of Neurosci., 1986, 6: 563-571.
McNaughton, B.L., Leonard, B., Chen, L. Cortical-hippocampal interactions and cognitive mapping: A hypothesis based on reintegration of the parietal and inferotemporal pathways for visual processing. Psychobiol., 1989, 17: 230-235.
Moore, J.W. Information processing in space-time by the hippocampus. Physiolog. Psychol., 1979, 7: 224-232.
Morris, R.G.M., Garrud, P., Rawlins, J.N.P. and O'Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature, 1982, 297: 681-683.
Morris, R.G.M., Schenk, F., Tweedie, F. and Jarrard, L.E. Ibotenate lesions of hippocampus and/or subiculum: Dissociating components of allocentric spatial learning. Eur.J. Neurosci. 1990, 2: 1016-1028.
Mutani, R. Cobalt experimental amygdaloid epilepsy in the cat. Epilepsia, 1967, 8, 1: 73-92.
Nachkebia, N.G. and Oniani, T.N. The role of the septo-hippocampal system in the regulation of learning and memory. In: Neurophysiology of Emotion, Memory and Sleep-Wakefulness Cycle. Metsniereba, Tbilisi, 1985: 59-99.
Nadel, L. Is the hippocampal formation preferentially involved in spatial behavior? Hippocampus, 1991, 1: 221-292.
Nadel, L. The psychobiology of spatial behavior: The hippocampal formation and spatial mapping. In: E. Alleva, H. P. Lipp, L. Nadel, A. Fasolo and L. Ricceri (Eds.). Behavioral Brain Research in Naturalistic and Semi-Naturalistic Settings: Possibilities and Perspectives. Kluwer Press, 1995.
Nyberg, L. McIntosh, A.R., Houle, S., Nilsson, L.G. and Tulving, E. Activation of medial temporal structures during episodic memory retrieval. Nature, 1996, 380: 715-717
O'Keffe, J. and Nadel, L. The Hippocampus as a Cognitive Map. Oxford. The Clarendon Press. 1978.
O'Reilly, R.C. and McClelland, J.L. Hippocampal conjunctive encoding, storage, and recall: avoiding a trade off. Hippocampus, 1994, 6: 661-682.
Olton, D.S. Mazes, maps and memory. Amer. Psychol., 1979, 34: 583-596.
Olton. D.S., Walker, J.A. and Gage, F.H. Hippocampal connections and spatial discrimination. Brain Res. 1978, 139: 295-308.
Oniani, T.N. The Integrative Function of the Limbic System. Metsniereba, Tbilisi, 1980 (in russ.)
Oniani, T.N. and Badridze J.K. About the behavioral correlates of the hippocampal theta rhythm. In T. Oniani (Ed) Neurophysiology of Emotion and Wakefulness-Sleep Cycle. Metsniereba, Tbilisi, 1974: 7-24 (in Russian).
Oniani, T.N. and Lortkipanidze, N.D. Influence of deprivation of paradoxical sleep on the learning and memory. In: T. Oniani (Ed.), Neurophysiology of Emotion, Memory and Sleep-Wakefulness Cycle. Tbilisi, Metsniereba, 1985: 199-214, (in Russian).
Oniani, T.N. and Nachkebia, A. Y. About the problem of hippocampo-mesodiencephalic relationships. Physiolog. J. of SSSR, 1982, 28: 684-693, (in Russian).
Oniani, T.N. and Nachkebia, N.G. The role of the septo-hippocampal system and paradoxical phase of sleep in the regulation of learning and memory. Neurobiology of Sleep-Wakefulness Cycle, 2001, 1(2): 64-94.
Papez, J.W. A proposed mechanism of emotion. Arch. Neurol. Psychiat. 1937, 38: 725-744.
Rempel-Clower, N.L., Zola, S.M., Squire, L.R., Amaral, D.G. Three cases of enduring memory impairment after bilateral damage limited to the hippocampal formation. J. Neurosci. 1996, 16: 5233-5255.
Roldan, E. The sleep cycle durating in normal and epileptic cats and humans. Activ. Nerv. Sup., 1969, 11, 2: 247-256.
Rudell, A. P., Fox, S. E., Ranck, I. B. Hippocampal excitability phaselocked to the theta rhythm in walking rats. Exp. Neurol., 1980, 68, 1: 87-96.
Sandkuchler, J., Maisch, B., Gebhart, G.F. and Zimmerman, M. reversible microblockade in spinal cord and brainstem of cats and rats by focal lidocaine. Pflug. Arch., 1995, 405: R41.
Scoville, W.B. and Milner, B. Loss of recent memory after bilateral hippocampal lesions. J. of Neurol. Neurosur., and Psych., 1957, 20: 11-21.
Shinkman, P.G. and Kaufman, K.P. Posstrial hippocampal stimulation and CER acquisition in the rat. J. of Comp. Physiol. Psychol. 1972, 80: 283-292.
Sideroff, S., Bueno, O.,Hirgch, A., Weyand, T., McGaugh, J.L. Retrograde amnesia initiated by low-level stimulation of hippocampal cytoarchitectonic areas. Exp. Neurol., 1974, 43: 285-297.
Simonov P.V. The role of hippocampus in the integrative activity of the brain. J. of High Nerv. Activ., 1972, 22: 1119-1124, (in Russian).
Solomon, P.R. Role of the hippocampus in the blocking and conditioned inhibition of the rabbit's nictating membrane response. J. of Comp. Physiol. Psychol. 1977, 91: 407-417.
Soumireu-Mourat B., Destrade, C. and Cardo, B. Effects of seizure and sub-seizure post-trial hippocampal stimulation on appetitive operant behavior in mice. Behav. Biol., 1975, 15: 303-316.
Squire, L.R. Memory and the hippocampus: A synthesis from findings with rats, monkeys, and humans. Psychol. Rev., 1992, 99: 195-231.
Squire, L.R., Ojemann, J.G., Meizin, F.M., Petersen S.E., Videen, T.O. and Raichle, M.E. Activation of the hippocampus in normal humans: A functional anatomical study of memory. Proc. Natl. Acad. Sci. USA, 1992, 89: 1837-1841.
Stein, D.G. and Chorover, S.L. Effects of post-trial electrical stimulation of hippocampus and caudate nucleus on maze learning in the rat. Physiol. and Behav., 1968, 3: 787-791.
Stern, C.E., Corkin, S., Gonzalez, R.I., Baker, J.R., Jenings, P.J., Carr, C.A., Sugiura, R.M.,Vedantham, V. and Rosen, B.R. The hippocampal formation participates in novel picture encoding: evidence from functional magnetic resonance imaging. Proc. Natl. Acad. Sci. USA, 1996, 93: 8660-8665.
Sutherland, R.J. and Rudy, J.W. Configural association theory: The role of the hippocampal formation in learning, memory and amnesia. Psychobiol., 1989, 17: 129-144.
Vardaris, R.M. and Schwartz, K.E. Retrograde amnesia for passive avoidance produced by stimulation of dorsal hippocampus. Physiol. and Behav., 1971, 6: 131-136.
Vinogradova, O.S. The Hippocampus and Memory. 1975. Moscow, Nauka, (in Russian).
Wyers, E.I., Peeke, H.V.S., Williston, I.S. and Here, M.I. Retroactive impairement of passive avoidance learning by stimulation of the caudate nucleus. Exp. Neurol., 1968, 22: 350-366.
Zhuravin, L.A., Bures, J. Extent of tetrodotoxin induced blockade examined by pupilary paralysis elicited by intracerebral injection of the drug. Exp. Brain Res., 1991, 83: 687-690.
Zola-Morgan, S. and Squire, L.R. Neuroanatomy of memory. Annu. Rev. Neurosci. 1993, 16: 547-563.
Zola-Morgan, S., Squire, L.R. and Amaral, D.G. Human amnesia and the medial temporal lobe region: Enduring memory impairment following a bilateral lesion limited to field CA1 of the hippocampus J. Neurosci. 1986, 6: 2950-2967.
Zornetzer, S.F., Boast, C. and Hamrick, M. Neuroanatomical localization and memory processing in mice: the role of the dentate gyrus of the hippocampus. Physiol. and Behav. 1974, 13: 569-575.