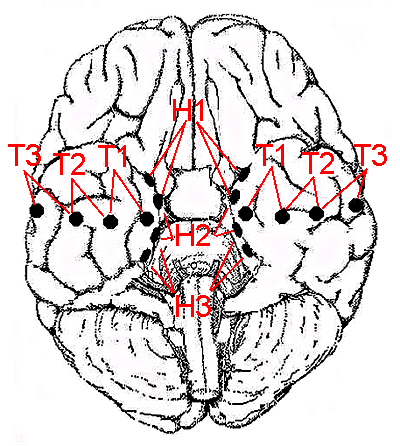
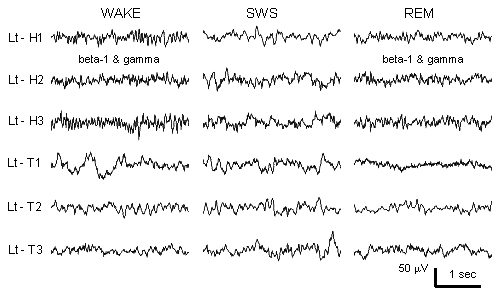
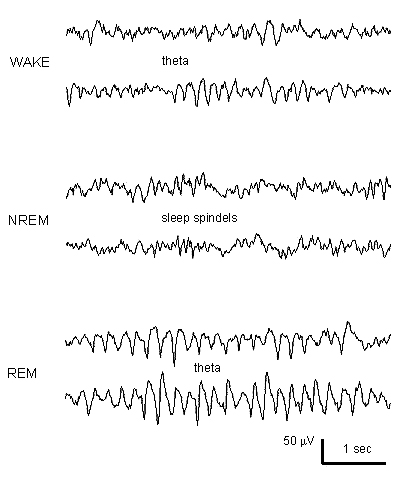
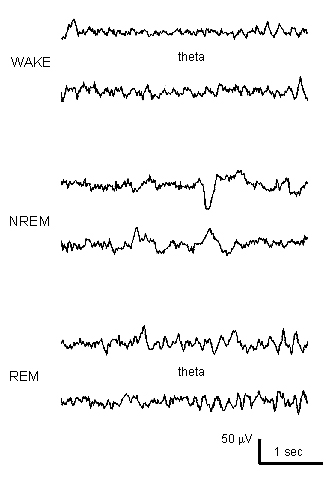
Arnolds, D.E.A.T., Lopes da Silva, F.H., Aitink, J.W., Kamp, A. and Boeijinga, P. The spectral properties of hippocampal EEG related to behaviour in man. Electroenceph. clin. Neurophysiol., 1980, 50: 324-328.
Bliss, T.V.P. and Collingridge, G.L. A synaptic medel of memory: long-term potentiation in the hippocampus. Nature, 1993, 361: 31-39.
Bliss, T.V.P. and Lomo, T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol., Lond., 1973, 232: 331-356.
Bragin, A., Jando, G., Nadasdy, Z., Hetke, J., Wise, K. and Buzsaki, G. Gamma (40-100 Hz) oscillation in the hippocampus of the behaving rat. J. Neurosci., 1995, 15: 47-60.
Bush, G., Luu, P. and Posner, M.I. Cognitive and emotional influence in anterior cingulate cortex. Trends in Cognitive Sciences, 2000, 4: 215-222.
Eichenbaum, H. A cortical-hippocampal system for declarative memory. Nature Reviews Neuroscience, 2000, 1: 41-50.
Fox, S.E. and Ranck, J.B. Electrophysiological characteristics of hippocampal complex-spike cells and theta cells. Experimental Brain Research., 1981.
Freemon, F.R. and Walter, R.D. Electrical activity of human limbic system during sleep. Comprehensive Psychiatry, 1970, 11: 544-551.
Gevins, A., Smith, M.E., McEvoy, L. and Yu, D. High-resolution EEG mapping of cortical activation related to working memory: effects of task difficulty, type of processing, and practice. Cereb. Cortex, 1997, 7: 374-385.
Giaquinto, S. Sleep recordings from limbic structure in man. Confin. Neurol., 1973, 35: 285-303.
Green, J.D. and Arduini, A. Hippocampal electrical activity in arousal. J. Neurophysiol., 1954, 17: 533-557.
Halgren, E., Babb, T.L. and Crandall, P.H. Human hippocampal formation EEG desynchronizes during attentiveness and movement. Electroenceph. clin. Neurophysiol., 1978, 44: 778-781.
Hirai, N., Uchida, S., Maehara, T., Okubo, Y. and Shimizu, H. Beta-1 (10-20 Hz) cortical oscillations observed in the human medial temporal lobe. Neuroreport, 1999, 10: 3055-3059.
Hirai, N., Uchida, S., Maehara, T., Okubo, Y. and Shimizu, H. Enhanced gamma (30-150 Hz) frequency in the human medial temporal lobe. Neuroscience, 1999, 90: 1149-1155.
Huerta, P.T. and Lisman, J.E. Heightened synaptic plasticity of hippocampal CA1 neurons during a cholinergically induced rhythmic state. Nature, 1993, 364: 723-725.
Ishihara, T. and N.Yoshii Multivariate analytic study of EEG and mental activity in juvenile delinquents. Electroenceph. clin. Neurophysiol., 1972, 33: 71-80.
Ishii, R., Shinosaki, K., Ukai, S., Inouye, T., Ishihara, T., Yoshimine, T., Hirabuki, N., Asada, H., Kihira, T., Robinson, S.E. and Takeda, M. Medial prefrontal cortex generates frontal midline theta rhythm. Neuroreport, 1999, 10: 675-679.
Jouvet, M., Michel, F. and Courjon, J. Sur un stade dactivite electrique cerebrale rapide au cours du sommeil physiologique. C. R. Soc. Biol., 1959, 153: 1024-1028.
Jung, R. and Kornmuller, A. Ein methodik der abteilung lokalsierter potential schwankingen aus subcorticalen hirnyebieten. Arch. Psychiat. Neuroenkr., 1938, 109: 1-30.
Kahana, M.J., Sekuler, R., Caplan, J.B., Kirschen, M. and Madsen, J.R. Human theta oscillations exhibit task dependence during virtual maze navigation. Nature, 1999, 399: 781-784.
Kemp, N. and Bashir, Z.I. Long-term depression: a cascade of induction and expression mechanisms. Progress in Neurobiology, 2001, 65: 339-365.
Konopacki, J., MacIver, M.B., Bland, B.H. and Roth, S.H. Carbachol-induced EEG "theta" activity in hippocampal brain slices. Brain Res., 1987, 405: 196-198.
Larson, J., Wong, D. and Lynch, G. Patterned stimulation at the theta frequency is optimal for the induction of hippocampal long-term potentiation. Brain Res., 1986, 368: 347-350.
Lisman, J.E. and Idiart, M.A.P. Storage of 7 plus-minus 2 short-term memories in oscillatory subcycles. Science, 1995, 267: 1512-1515.
Maguire, E.A., Burgess, N. and O'Keefe, J. Human spatial navigation: cognitive maps, sexual dimorphism, and neural substrates. Current Opinion in Neurobiology, 1999, 2: 171-177.
Meador, K.J., Thompson, J.L., Loring, D.W., Murro, A.M., King, D.W., Gallagher, B.B., Lee, G.P., Smith, J.R. and Flanigin, H.F. Behavioral state-specific changes in human hippocampal theta activity. Neurology, 1991, 41: 869-872.
O'Keefe, J. and Burgess, N. Theta activity, virtual navigation and the human hippocampus. Trends in Cognitive Science, 1999, 3: 403-406.
O'Keefe, J. and Dostrovsky, J. The hippocampus as a spatial map: preliminary evidence from unit activity in the freely moving rat. Brain Res., 1971, 34: 171-175.
Poe, G.R., Nitz, D.A., McNaughton, B.L. and Barnes, C.A. Experience-dependent phase-reversal of hippocampal neuron firing during REM sleep. Brain Research, 2000, 855: 176-180.
Sano, K., Mayanagi, Y., Sekino, H., Ogashiwa, M. and Ishijima, B. Results of stimulation and destruction of the posterior hypothalamus in man. J Neurosurg, 1970, 33: 689-707.
Scoville, W.B. and Milner, B. Loss of recent memory after bilateral hippocampal lesions. Journal of Neurology, Neurosurgery and Psychiatry, 1957, 20: 11-21.
Shimazono, Y., Horie, T., Yanagisawa, N., Hori, N., Chikazawa, S. and Shozuka, K. The correlation of the rhythmic waves of the hippocampus with the behaviors of dogs. Neurol. med. -chirur., 1960, 2: 82-88.
Shimizu, H., Suzuki, I., Ohta, Y. and Ishijima, B. Mesial temporal subdural electrode as a substitute for depth electrode. Surg. Neurol., 1992, 38: 186-191.
Steward, M., Luo, Y. and Fox, S.E. Effects of atropine on hippocampal theta cells and complex-spike cells. Brain Res., 1992, 591: 122-128.
Suzuki, W.A. and Clayton, N.S. The hippocampus and memory: a comparative and ethological perspective. Current Opinion in Neurobiology, 2000, 10: 768-773.
Tesche, C.D. Non-invasive detection of ongoing neuronal population activity in normal human hippocampus. Brain Res., 1997, 749: 53-60.
Uchida, S., Maehara, T., Hirai, N., Kawai, K. and Shimizu, H. Theta oscillation in the anterior cingulate and beta-1 oscillation in the medial temporal cortices: A human case report. (Submitted).
Uchida, S., Maehara, T., Hirai, N., Okubo, Y. and Shimizu, H. Cortical oscillations in human medial temporal lobe during wakefulness and all-night sleep. Brain Res., 2001, 891: 7-19.
Vertes, R.P., Albo, Z. and Viana di Prisco, G. Theta-rhythmically firing neurons in the anterior thalamus: Implications for mnemonic functions of Papez's circuit. Neuroscience, 2001, 104: 619-625.